3月15日,plant physiology在线发表了华中农业大学宋波涛教授团队题为“tcp transcription factor stast1 represses potato tuberization by regulating tuberigen complex activity”的研究成果,该文章发现了一个新的tcp转录因子stast1,可以抑制马铃薯块茎的形成,stast1还可以通过激活马铃薯ga20氧化酶基因(stga20ox1)的表达,调控ga应答。结果表明,stast1通过调节植物激素水平发挥块茎形成抑制因子的作用。迈维代谢为其提供了植物激素检测与分析服务。

新型器官的出现是植物适应环境做出的改变,可确保植物能够应对不利条件或在极端环境下生存。马铃薯通过地下分枝或匍匐茎分化成块茎而形成块茎,这使得植物能够在寒冷的冬季生存。马铃薯是研究地下特化器官形成的模型系统。了解马铃薯植物的块茎化不仅有助于在气候变化条件下保证马铃薯的生产,而且有助于更好地了解其他植物的储存器官形成。与开花相似,匍匐茎表达的开花位点t-like (ft-like)蛋白self-pruning 6a (stsp6a)通过与bzip转录因子stabi5-like 1 (stabl1)和stfd-like 1 (stfdl1)结合,在匍匐茎近顶端引起转录重编程,从而在块茎形成过程中发挥重要作用。然而,调节广泛保守的ft-bzip相互作用的分子机制在很大程度上仍未被探索。
stast1基因的沉默导致块茎的形成和生命周期的缩短
为了研究 stast1 在块茎化过程中的基因功能,作者生成了stast1 过表达植株oe-stast1和stast1的 rna 干扰植株ri-stast1。4周龄的植株,stast1沉默植株在 5 d 内结块,野生型和过表达植株结块较晚。生长 120 d 后,stast1沉默植株块茎数量明显少于过表达和野生植株。与野生型相比,stast1沉默 和stast1过表达 植物分别表现出更早和更晚的叶片衰老(图 1),植物的生命周期分别缩短和延长(图1)。综上所述,这些发现表明 stast1 负向调控块茎化时间和植株成熟度。作者又评估了新收获块茎的休眠表型。 发现,stast1过表达块茎释放休眠的时间最长,其次是野生型,stast1沉默植株萌芽时间最短。这表明 stast1 对块茎萌发也有负调控作用。
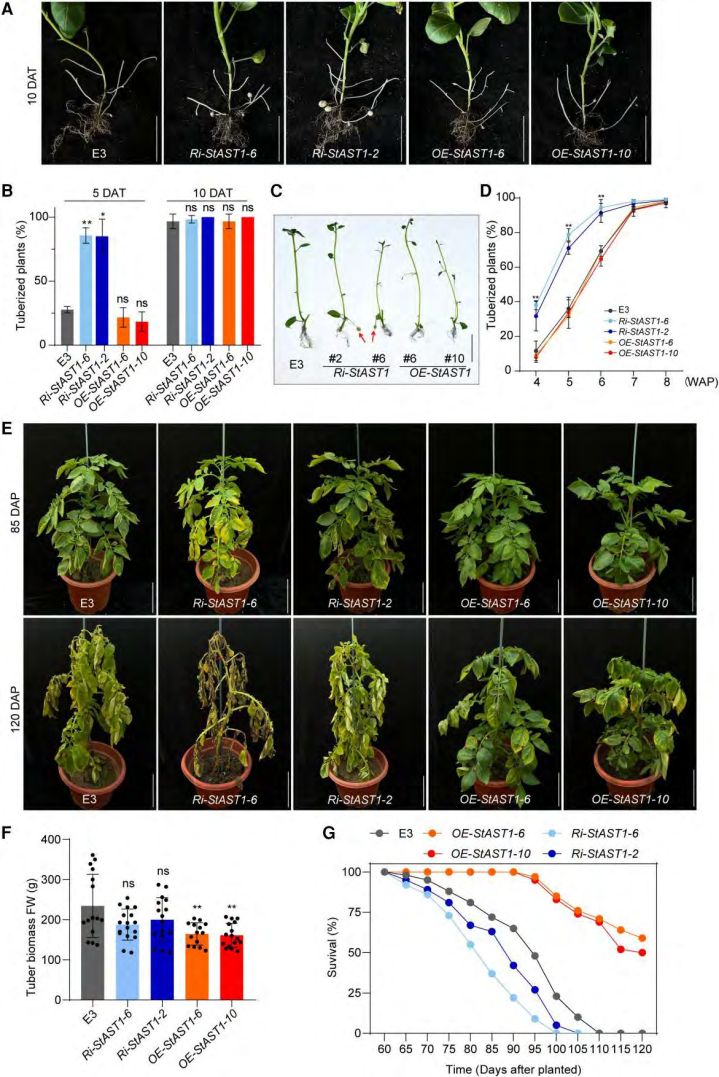
图1 stast1转化材料的块茎化和成熟表型
stast1激活stga20ox1调控马铃薯的ga反应
为了确定stast1在早期块茎化的转录变化,作者对ri-stast1 植株和野生型材料进行转录组测序。ri-stast1 植物中发现1,906个差异表达基因,在激素调节途径中,degs 主要富集在 aba 途径中,表明干扰 stast1 可能会影响植物对 aba 的响应。体外施用 5 μm aba 能显著地促进 ri-stast1植株提前结块(图2)除此之外,与块茎功能相关的基因,如stpp2c21 和 atsps3f-like,糖外流转运体 stsweet12e 和 stsweet11 在沉默stast1植株中上调,这些数据共同表明,干扰 stast1 会导致多个具有促进块茎化功能的基因上调。
在下调表达基因中,stga20ox1和 stga20ox3 在沉默植株中显著下调。ga20ox 编码一种参与ga生物合成的关键酶,它催化产生具有生物活性的 ga 的直接前体。沉默 stga20ox1 可促进块茎的提前形成。为了研究 stast1 对 ga 合成途径中代谢物的影响,收集了野生型和ri-stast1 的匍匐茎尖,并用于 ga 含量的测定。 检测到了 18 种代谢物中的 16 种。在 ri-stast1 的匍匐茎中,由 stga20ox 合成的 ga20、ga9 和 ga19 的含量显著降低(图2)。相比之下,具有生物活性的 ga(如 ga1)则有所增加,而 ga3 则没有明显变化(图 2)。 这些结果表明,stast1 通过影响 ga 合成途径在块茎形成过程中发挥作用。 为了测试 ga 的反应,在块茎诱导培养基下对 ri-stast1 植株施加 ga3,所有 ri-stast1 植株都产生了块茎。表明干扰 stast1 会导致对 ga 的敏感性降低。后续,作者通过双荧光素酶报告实验和emsa证明stast1 能特异性地与 stga20ox1 结合并激活它。
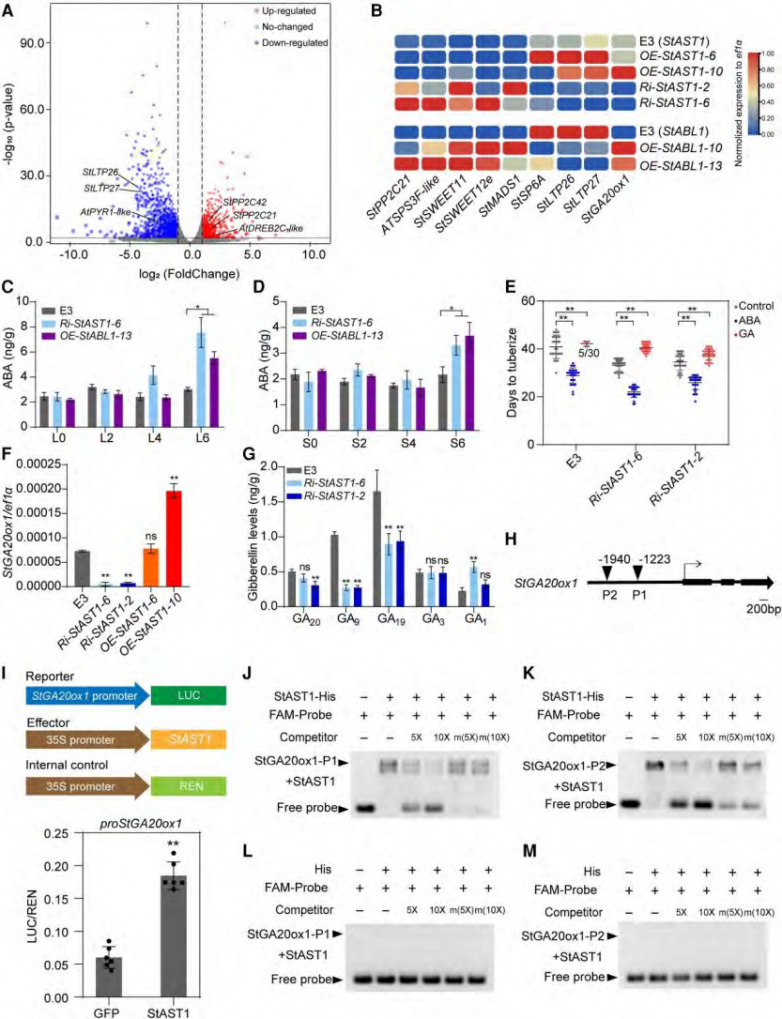
图2 stast1敲除对马铃薯块茎转录变化
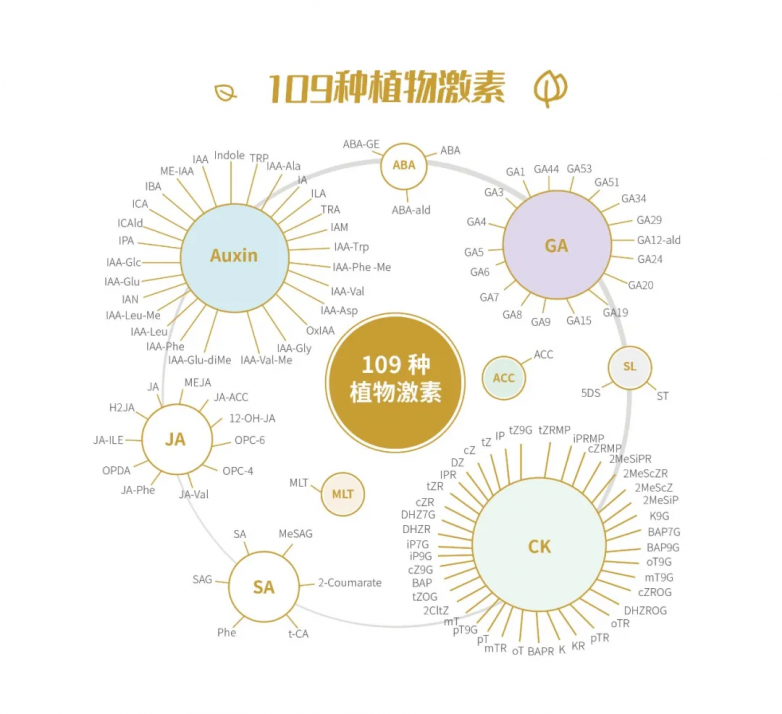
迈维代谢开发了基于lc-ms/ms技术的植物激素全家桶产品,包括9大类109种植物激素,其中生长素类物质包含27种,联合外标 内标实现检测物质的绝对定量。欢迎垂询!
免责声明:市场有风险,选择需谨慎!此文仅供参考,不作买卖依据。